

Parasites which reside within the blood or internal organs of the host have logistical problems in terms of infecting a new host. In contrast to fecal-oral transmission, where infective stages are excreted into the environment, potential new hosts would not normally come into contact with the parasite. (In evolutionary terms, transmission by blood transfusion would be a very recent event.) Predator-prey transmission is one strategy used by protozoa such as Toxoplasma and Sarcocystis to overcome these transmission barriers. As the name implies, predator-prey transmission involves two distinct hosts. The predator acquires the infection by eating an infected prey. This will induce an intestinal infection in the predator and result in the excretion of infective stages in the feces. The prey, generally a herbivore, will become infected by eating the infective stages it encounters in the environment. Following ingestion by an appropriate prey the parasite will cross the intestinal epithelium and infect internal organs or tissues within the host, where it waits for the next predator to ingest its prey. [Page contents]
Vector transmission is the other strategy used by protozoan parasites which inhabit the blood or internal tissues within its host. This strategy involves a hematophogous (ie, blood feeding) arthropod serving as an intermediary between successive vertebrate hosts. Several human diseases caused by protozoa are transmitted by a variety of arthropod vectors (Table). The vectors are not simply 'flying syringes', but represent a second host for the protozoan parasite. Thus the life cycle of vector transmitted diseases also involves complex interactions protozoan-vector interaction analogous to the complex human-protozoan interactions. Vector transmission probably evolved multiple times.
| Vector Transmitted Protozoa | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
||||||||||||||||||||||||||||
Vector transmission also involves complex interactions between humans and vectors. This includes the biology of human-arthropod interactions, as well as ecological considerations. Thus vector transmitted parasites exhibit complex life cycles involving interactions between humans, protozoa, and arthropods. The biology of vectors and their interactions with humans provide possible means for controlling the transmission of these diseases.
|
|
|
|
 |
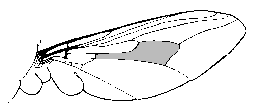 |
The genus Glossina, common name tsetse, contains at least 30 species or subspecies of flies found in subsaharan Africa. Adult tsetse are yellow, brown, or black flies 6-14 mm long. A diagnostic character of the genus is the 'hatchet cell' found in the center of the wings (Figure). Most species fall into one of two major groups represented by G. palpalis and G. morsitans. The palpalis group is associated with riverine ecologies and is frequently found near streams, rivers, and lakes in west and central Africa. The morsitans group is most frequently associated with savannah woodlands and dry bush country in east Africa. The palpalis and morsitans groups are associated with the transmission of Trypanosoma gambiense and T. rhodesiense, respectively. The differences in the ecologies and interactions with reserviors of these two types of tsetse contribute the different manifestations of diseases caused by the two African trypanosme species (See table in notes on African trypanosomes.)
Both the male and female tsetse are voracious feeders and the important vector species tend to feed on large mammals. Tsetse means 'fly destructive to cattle' in Sechuana (hence tsetse 'fly' is redundant). The bite of the tsetse is painful, but usually of minor consequence. A welt will occasionally form and some individuals will become sensitive to the saliva. However, tsetse are persistent feeders and can be quite aggravating to humans and animals. Tsetse are 'pool' feeders (ie, telmophages) which lacerate the skin with their mouth parts and then ingest the blood and lymph which flow into the superficial lesion. Trypanosome transmission occurs via the saliva which is injected into the bite site. The saliva contains substances which dilate blood vessels and prevent coagulation.
The life cycle of African trypanosomes involves alternating between the vertebrate host and the tsetse vector. Trypomastigotes within the blood stream of the vertebrate host are ingested by the tsetse and convert to procyclic trypomastigotes which replicate within the gut of the tsetse. To complete the life cycle, these procyclic forms must make their way to the saliva glands. The exact mechanism by which the parasite migrates from the tsetse gut to the saliva glands is not known. Two routes have been proposed: 1) the classical route in which the parasite 'backtracks' through the digestive system and migrates up the salivary duct, or 2) the direct route in which the parasite penetrates the peritrophic membrane and gut epithelium to gain access to the hemolymph. Either route presents barriers and illustrates the complex interactions between the protozoan parasite and vector (Figure).
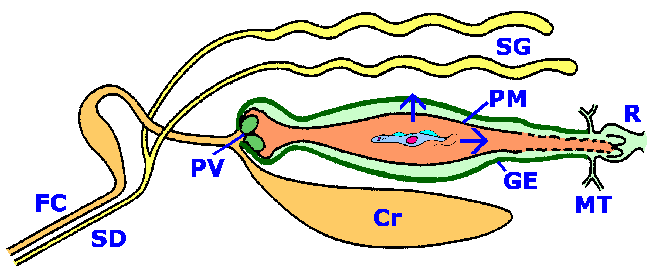 |
| Schematic representation of trypanosome-tsetse interactions. Trypomasitgotes are taken up with the blood meal and pass through the food canal (FC) and crop (Cr). The blood meal is encased in the peritrophic membrane (PM) by the proventricular valve (PV) as it enters the midgut. Migration of the procyclic trypomastigotes from the midgut to the salivary glands (SG) involves two possible routes (depicted by arrows): the 'classical' and the 'short circuit'. The classical route invovles a migration towards the hindgut (Malpighian tubes, MT and rectum, R) and exiting the peritrophic membrane. The procyclic then migrates forward through the ectoperitrophic space, exits at the proventricular valve, passes up the food canal, and enters the salivary duct (SD) to gain access to the salivary glands.The short circuit route involves a direct penetration of the peritrophic membrane and gut epithelium (GE) to gain access to the hemocoel. From the hemocoel the trypanosomes find and penetrate the salivary glands. |
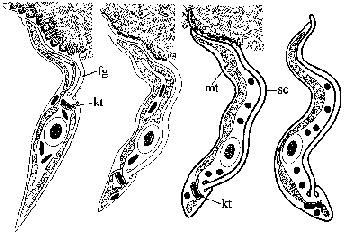 |
| Diagram of developmental stages found in the tsetse salivary gland. From Tetley and Vickerman (1985) J. Cell Sci. 74:1. |
The parasite converts to an epimastigote form after reaching the salivary gland. The epimasigote attaches to the epithelial cells of the salivary glands via its flagellum (fg) and undergoes further replication. Differentiation into metacyclic trypomastigotes involves the appearance of the surface coat (sc) and changes in the mitochondria (mt) to accompany the migration of the kinetoplast (kt) to the posterior end (Figure). These developmental changes are necessary preparations for becoming infective to the mammalian host. The mature metacyclic trypomastigote detatches from the epithelial cells and remains in the lumen of the salivary gland until being expelled during tsetse feeding.
(Return to African Trypanosome Life Cycle or go to Control Strategies.)
|
|
|
|
 |
| Picture modified from A.M. Rose (http://www.arose.net/triatoma/pics.htm) and used with permission (© H Knüttel & AM Rose). |
Several genera and species of triatomines are capable of transmitting Trypanosoma cruzi. The most important vectors are: Triatoma infestans, Panstrongylus megistas, and Rhodnius prolixus. Triatomines are generally large insects ranging in size from 5-30 mm. They are primarily New World insects and range from Argentina through the United States. The life cycle consists of five nymphal instars followed by sexually mature adults. Only the adults have wings. Triatomines feed on blood through out their life and all stages can become infected with T. cruzi. Common names include assassin bug, kissing bug (in reference for its tendency to bite around the face) and conenose bug (in reference to their pointed heads).
Most triatomines are not host specific and feed on a wide range of mammals as well as birds and reptiles. Transmission of T. cruzi is often associated with domiciliary species which have adapted to living in human dwellings. Such dwellings are generally adobe or thatched roof huts which provide plenty of hiding places for the bugs. The bugs come out at night to feed upon sleeping people. They are capillary feeders and feeding times range from 3-30 minutes. Adults can can up to 0.25 ml of blood per feeding and a total of 4-10 ml of vertebrate blood may be consumed by a triatomine bug during its lifetime. The bites are usually painless, despite the large proboscis. It is assumed that the saliva contains an anesthesia. Although triatomine feeding causes little pain, their bites can cause an uncomfortable lesion at the site of feeding and can elicit a hypersensitivity reaction.
Triatomines become infected when bloodstream trypomastigotes are ingested with the blood meal. The trypomasigotes convert to epimastigotes which are the replicative form found primarily in the midgut. The epimastigotes will develop into non-dividing metacyclic trypomasitigotes. Metacyclogenesis is almost exclusive to the rectum and correlates with attachment to the rectal epithelium. Vector-parasite interactions are important for the successful replication and differentiation of T. cruzi.(See also: Kollien and Schaub, The development of Trypanosoma cruzi in triatominea. Parasitol. Today 16:381; 2000.) Infectious metacyclic trypomastigotes are excreted with the feces. This type of transmission is referred to as 'hindgut station' or 'stercorarian' and is considerably less efficient than salivarian transmission.
(Go to T. cruzi Life Cycle, Ecology of Transmission, or Control Strategies.)
|
|
|
|
 |
| From Peter and Gilles (Color Atlas of Tropical Medicine and Parasitology) |
Sand flies are the vectors for Leishmania as well as some bacterial (bartonellosis) and viral (sand fly fever) diseases. Adult sand flies are about 2 mm in length and characterized by hair bodies and wings (Figure?). Genera of sand flies fall into two groups: Phlebotomus and Sergentomyia found in the Eastern hemisphere (Europe, Asia, Africa, Australia), and Lutzomyia, Brumptomyia and Warileya in the Western hemisphere (ie, New World). In general, the Old World sand fly species live in desert or semi-arid ecosystems and the New World species are forest dwelling. Some of the Old World species will breed in peridomestic situations and enter human habitations, whereas disease transmission in the New World is associated with humans living or working near the forest.
The sand flies responsible for the transmission of leishmaniasis are Phlebotomus and Lutzomyia in the Old and New Worlds, respectively. The spatial distribution of leishmaniasis and other sand fly transmitted diseases tends to be patchy due to the limit flying range of the sand fly. Their flying is often described as 'hopping', characterized by short bursts of flights separated by a couple of seconds of rest. Sand flies have a short flight range and are usually found within a dozen meters of a breeding site.
As a general rule only female sandflies take blood meals and they feed most actively at twilight or night, or in low-light conditions such as shade. They have short mouthparts and are pool feeders. The bite produces a rose-colored papule surrounded by a erythematous area about 10-20 mm in diameter.
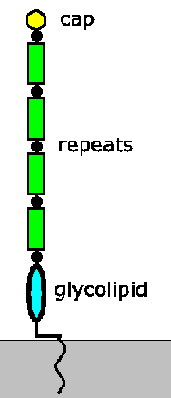 Amastigote-infected
macrophages are ingested with the blood meal and transform into promastigotes
(see Leishmania Life Cycle).
These procyclic promastigotes attached to the midgut epithelium of the fly and
replicate. Attachment is mediated by a lipophosphoglycan (LPG). LPG is an abundant
cell surface associated glycoconjugate composed of three major domains: glycolipid
membrane anchor, disaccharide repeats, and a small cap (Figure). The LPG undergoes
biochemical changes as the parasites mature into metacyclic promastigotes. The
disaccharide repeats approximately double in number leading to an elongation
of the LPG and the cap changes from galactose to
arabinose. Changing the cap structure is believed
to be associated with detachment from the gut epithelium in that adherence is
believed to be associated with galactose specific lectins found in the sandfly's
gut epithelium. The elongaton of LPG is associated with increased complement
resistance suggesting additional roles for LPG in terms of infectivity to the
vertebrate host. (For a review on Leishmania glycoconjugates see: Descoteaux
and Turco, Biochem.
Biophys. Acta 1455:341; 1999.)
Amastigote-infected
macrophages are ingested with the blood meal and transform into promastigotes
(see Leishmania Life Cycle).
These procyclic promastigotes attached to the midgut epithelium of the fly and
replicate. Attachment is mediated by a lipophosphoglycan (LPG). LPG is an abundant
cell surface associated glycoconjugate composed of three major domains: glycolipid
membrane anchor, disaccharide repeats, and a small cap (Figure). The LPG undergoes
biochemical changes as the parasites mature into metacyclic promastigotes. The
disaccharide repeats approximately double in number leading to an elongation
of the LPG and the cap changes from galactose to
arabinose. Changing the cap structure is believed
to be associated with detachment from the gut epithelium in that adherence is
believed to be associated with galactose specific lectins found in the sandfly's
gut epithelium. The elongaton of LPG is associated with increased complement
resistance suggesting additional roles for LPG in terms of infectivity to the
vertebrate host. (For a review on Leishmania glycoconjugates see: Descoteaux
and Turco, Biochem.
Biophys. Acta 1455:341; 1999.)
There are also factors within the sandfly saliva which potentiate the infectivity of Leishmania for the vertebrate host. Some of these salivary gland compounds have immunosuppressive activities against lymphocytes and macrophages which may explain the potentiation.
(Return to Leishmania Life Cycle or go to Control Strategies.)
|
|
|
|

|
|
|
|
|
|
|
|
See disease specific control activities:
These pages are developed and maintained by Mark F. Wiser, Tulane University (© 2001). Last update on February 11, 2022 .